Plattform für Wasser, Gas und Wärme
 zum Hypolimnion (Abbau des organischen Materials und Sauerstoffzehrung).
In Rot die wichtigsten Prozesse: PP: Primärproduktion; R: Respiration; NEP: Netto-Ökosystemproduktion; WCM: Abbau in der Wassersäule;
SOU: Sauerstoffaufnahme ins Sediment; NS: Netto-Sedimentation; RED: Fluss reduzierter Substanzen aus dem Sediment; GA: Gasaustausch.
In Grün die verschiedenen Formen von Kohlenstoff: POC: partikulärer organischer Kohlenstoff; CO2; CH4; CaCO3: anorganischer Kohlenstoff.")
 und Netto-Sedimentation (NS) im Hypolimnion für die drei wichtigsten Seetypen. Zwei Faktoren tragen zur Unsicherheit der Kohlenstoffbilanz im Hypolimnion bei: allochthon (see-extern) eingetragenes terrestrisches Material, das die NS erhöht, und die eutrophe Vergangenheit, die den Fluss von reduzierten Substanzen (RED) erhöht. Entsprechend müssen diese Beiträge beim Typ 1 (RED-Altlast) und Typ 2 (NS-allochthon) ausgeschlossen werden. Die drei mit * markierten Anteile der aktuellen NEP sind nicht zweifelsfrei bestimmbar und werden daher vernachlässigt. Die NEP wird in diesen Fällen leicht unterschätzt.")
 O2-Gehalt im gesamten Genfersee (in Mio t; hellgrün) und O2-Gehalt im Hypolimnion (dunkelgrün) für 1980 bis 2016.
b) Verlauf der volumengewichteten, mittleren O2-Konzentrationen im Hypolimnion für zwei sehr verschiedene Sommer. Ausgewählte Start- und Enddaten sind mit einem Kreis gekennzeichnet.")
![Fig. 4 a) Zeitreihen der AHM (ausgedrückt als Sauerstoffverbrauch pro Tag während der produktiven Saison) seit 1957. Grau: der gleitende Mittelwert über jeweils zehn Jahre. Durch die Grösse des Genfersees haben vertikale Oszillationen der Wasserschichten eine starke Auswirkung auf die saisonale O2-Bilanz, was zu methodischen Fehlern und deshalb zu grossen jährlichen Unterschieden in der berechneten AHM führt.
b) NEP (in g C m–2 [180 Tage]–1) abgeschätzt aus AHM plus NS für jede produktive Saison. Grau: der gleitende Mittelwert über jeweils zehn Jahre.
c) Mittlere P-Konzentrationen (TP und PO4) im ganzen See und Zielwert 2020 [15, 17].](/media/7564/kiefer-4.bmp?width=760&height=440&bgcolor=e8e8e8 "Fig. 4 a) Zeitreihen der AHM (ausgedrückt als Sauerstoffverbrauch pro Tag während der produktiven Saison) seit 1957. Grau: der gleitende Mittelwert über jeweils zehn Jahre. Durch die Grösse des Genfersees haben vertikale Oszillationen der Wasserschichten eine starke Auswirkung auf die saisonale O2-Bilanz, was zu methodischen Fehlern und deshalb zu grossen jährlichen Unterschieden in der berechneten AHM führt.
b) NEP (in g C m–2 [180 Tage]–1) abgeschätzt aus AHM plus NS für jede produktive Saison. Grau: der gleitende Mittelwert über jeweils zehn Jahre.
c) Mittlere P-Konzentrationen (TP und PO4) im ganzen See und Zielwert 2020 [15, 17].")
 Zusammensetzung der Kohlenstoffsenken im Baldeggersee (BAL), Brienzersee (BRI) und Genfersee (GEN), als drei Vertreter der Seetypen 1 bis 3 (Fig. 2).
b) Beispiel eines terrestrischen Eintrags: Schwemmgut im Brienzersee nach extremem Hochwasser im August 2005. (Foto: Arnold Gertsch im August 2005)")
![Fig. 6 Für elf Seen berechnete aktuelle NEP (in g C m–2 pro 180 Tage Schichtungsdauer) abgestuft nach Intensität der grünen Farbskala (Daten aus [9]).
In der Arbeitshilfe [1] sind die vorhandenen NEP-relevanten Daten und die Typisierung der Schweizer Seen (inkl. der drei Grenzseen) zusammengestellt.](/media/7566/kiefer-6.bmp?width=760&height=440&bgcolor=e8e8e8 "Fig. 6 Für elf Seen berechnete aktuelle NEP (in g C m–2 pro 180 Tage Schichtungsdauer) abgestuft nach Intensität der grünen Farbskala (Daten aus [9]).
In der Arbeitshilfe [1] sind die vorhandenen NEP-relevanten Daten und die Typisierung der Schweizer Seen (inkl. der drei Grenzseen) zusammengestellt.")
Mit weit über 1000 Seen ist die Schweiz ein seenreiches Alpenland. Das Bundesgesetz über den Schutz der Gewässer (GSchG, SR 814.20) sowie die Gewässerschutzverordnung (GSchV, SR 814.201) haben zum Ziel, eine gute Qualität der Schweizer Gewässer sicherzustellen, sie vor nachteiligen Einwirkungen zu schützen und ihre nachhaltige Nutzung zu gewährleisten (Art. 1 GSchV). Dank Grenzwerten in den Zuflüssen haben die Nährstoffkonzentrationen in fast allen Seen in den letzten Jahrzehnten deutlich abgenommen. Jedoch ist die Sauerstoffzehrung im Hypolimnion (geschichtetes Tiefenwasser) vieler Seen nach wie vor hoch. Die Anforderung, dass Seewasser zum Schutz höherer Lebewesen zu jeder Zeit und in jeder Tiefe mindestens 4 mg l–1 Sauerstoff (O2) enthalten soll, wird deshalb auch heute oft nicht eingehalten. Es stellt sich daher die Frage, ob die Primärproduktion seit der Eutrophierungsphase teilweise noch gar nicht abgenommen hat.
Die GSchV (Anhang 2, Ziffer 13) gibt eine klare Zielsetzung vor: Der Nährstoffgehalt in einem See soll höchstens eine «mittlere Produktion» von Biomasse zulassen. Die Quantifizierung dieser zeitlich stark variierenden Primärproduktion (PP) durch Fotosynthese von Algen und Cyanobakterien ist jedoch alles andere als einfach und beschäftigt die Wissenschaft seit Jahrzehnten (s. Box). Die Kenntnis darüber, wie viel organisches Material ein See produziert, ist aus mehreren Gründen von Bedeutung:
– Das pflanzliche Plankton bildet die Basis der Nahrungskette eines Sees und hat deshalb einen vielfältigen Einfluss auf alle Lebewesen (Zooplankton, Fische etc.) und somit auch auf die Biodiversität eines Sees.
– Eine übermässige Primärproduktion hat – besonders bei flachen Seen – negative Auswirkungen im Tiefenwasser. Sie verursacht durch Zehrung von O2 lebensfeindliche Bedingungen für höhere Lebewesen und schränkt deshalb den Lebensraum für (kälteliebende) Fische ein. Die Aufbereitung von Trinkwasser wird bei anoxischen Bedingungen wegen der Bildung von reduziertem Eisen und Mangan sowie Geruchsstoffen technisch deutlich aufwendiger.
– Seen sind Naherholungs- und Lebensraum. Daher soll auch aus gesundheitlichen und ästhetischen Gründen eine übermässige Produktion von Algen und Cyanobakterien vermieden werden.
– Seen sind ein wichtiger Teil des Kohlenstoffkreislaufs der Erde. Anorganischer Kohlenstoff (C) in Form von CO2 aus der Atmosphäre wird von den Algen in Biomasse umgewandelt. Seesedimente, in denen absinkendes nicht-abgebautes organisches Material eingelagert wird, sind wichtige C-Speicher.
Die Intensität der PP wirkt sich im gesamten Seevolumen aus. Die PP erfolgt im Epilimnion (Oberflächenschicht), weil hier Nährstoffe und Licht vorhanden sind, die für den Aufbau der Biomasse benötigt werden. Zooplankton bedient sich an dieser Biomasse, Bakterien bauen einen Teil des organischen Materials wieder ab und machen die Nährstoffe erneut verfügbar. Der Anteil des organischen Materials, der schliesslich ins Hypolimnion absinkt, wird als Netto-Ökosystemproduktion (engl: Net Ecosystem Production, NEP) definiert (Box). Es ist diese NEP, welche die O2-Zehrung im Hypolimnion der meisten Schweizer Seen prägt.
In diesem Artikel wird aufgezeigt, wie die NEP mit vergleichsweise wenig Aufwand aus Monitoring-Daten bestimmt werden kann. Die NEP stellt ein robustes Mass für die Entwicklung der Trophie, des Phosphorhaushalts und der Sauerstoffzehrung eines Sees dar und unterliegt weniger starken Schwankungen als die PP, die nur ungenau und mit grossem Aufwand erfassbar ist. Momentaufnahmen der PP werden heute nur noch im Genfersee und in den beiden Becken des Luganersees gemessen. Die PP kann sehr kurzfristig – z. B. beim Vorbeiziehen einer Wolke – variieren. Monatliche oder zweiwöchentliche Messungen der PP müssen daher sorgfältig auf ein ganzes Jahr hochgerechnet werden und erlauben erst über lange Zeiträume eine sinnvolle Interpretation. Auch der Versuch, die PP mittels Chlorophyll-a-Gehalt zu erfassen, ergibt nicht immer ein korrektes Abbild.
Im Folgenden wird dargestellt, wie aus kontinuierlichen Profilen der Nährstoffkonzentrationen von Phosphor (P) und Stickstoff (N) sowie des O2-Gehalts die NEP bestimmt werden kann. Diese Parameter werden üblicherweise in verschiedenen Schweizer Seen beim regulären Monitoring gemessen. Damit bietet sich die Möglichkeit, die Entwicklung der NEP und somit der Trophie eines Sees zeitlich zu verfolgen.
Netto-Ökosystemproduktion (NEP)
Primärproduktion (PP) ist die Assimilation von Kohlenstoff (C) aus der Atmosphäre mittels Sonnenlicht durch Algen und Cyanobakterien, welche am Anfang der Nahrungskette stehen. Die Netto-Ökosystemproduktion (NEP) ist jene Menge der PP, die aus dem Oberflächen- ins Tiefenwasser absinkt, nachdem schon in der produktiven Zone ein Teil durch Respiration (R) wieder abgebaut worden ist.
Netto-Ökosystemproduktion (NEP) = Primärproduktion (PP) – Respiration (R)
Die Tiefe, die man als Übergang zwischen Oberflächen- und Tiefenwasser festlegt, hat einen Einfluss auf die NEP, da auch in der produzierenden Schicht schon organisches Material abgebaut wird. Die sogenannte Kompensationstiefe kann zur Unterscheidung verwendet werden. Sie ist jene Tiefe, in der PP und R entgegengesetzt gleich gross sind. In diesem Artikel wurde zur Berechnung der NEP die Bilanz für den Genfersee in 15 m Tiefe festgelegt (entspricht in etwa dem Epilimnion, basierend auf Temperaturprofilen). Wichtig für die Vergleichbarkeit ist, dass für einen bestimmten See von Jahr zu Jahr die gleiche Tiefe verwendet wird. Für die Bestimmung der PP (tiefenabhängig oder integriert über die produzierende Schicht) gibt es verschiedene Methoden, die nachfolgend vorgestellt werden. R im Epilimnion abzuschätzen wäre extrem aufwendig und wurde bisher nicht realisiert. Die Berechnung der NEP mit dieser Formel ist somit nicht einfach möglich
Für die praktische Bestimmung von PP und NEP werden verschiedene Methoden verwendet, die hier nur kurz erwähnt werden, jedoch in der Arbeitshilfe [1] eingehender beschrieben sind.1 Bei der 14C-Methode werden Wasserproben in Glasflaschen mit markiertem Bikarbonat während einiger Stunden im See inkubiert und danach die assimilierte Menge 14C bestimmt [2]. Die 13C-Methode macht sich die ungleiche Assimilationsgeschwindigkeit der natürlichen Isotope 12C und 13C zunutze, aus deren unterschiedlichem Verhältnis im Wasser bzw. Plankton auf die Fotosyntheserate geschlossen werden kann [3]. Da bei der Fotosynthese O2 gebildet wird, kann mit der Tagesgang-Sauerstoffkurve (Diel-Methode) aus der integralen Tagesproduktion und Nachtkonsumation PP und R und aus deren Differenz die NEP berechnet werden [4]. Grosses Entwicklungspotenzial mit ersten Anwendungen hat die Nutzung von Fernerkundungsdaten der neuesten Sentinel-Satelliten. Sie erlauben die spektralanalytische Messung der Pigmentabsorption von Algen mit einer Auflösung von aktuell 300 m [5], woraus mit Modellen die PP bestimmt wird [6].
Beim Wachsen der Algen in der produktiven Zone und bei deren Export ins Hypolimnion wird auch eine zur NEP proportionale Menge P exportiert. Es ist daher möglich, anhand eines saisonalen Nährstoffbudgets des Epilimnions, z. B. P und N, die NEP zu berechnen. Dazu müssen auch die über den Sommer ein- und ausgetragenen Frachten bekannt sein sowie das Verhältnis C : P bzw. C : N, in welchem die Algen und Cyanobakterien die Nährstoffe einbauen. Anhand dieser Daten, der Dauer der produktiven Saison und der Hypolimnionfläche kann die durchschnittliche NEP pro m2 und Zeit berechnet werden. Allerdings sind zusätzlich zu den regelmässigen Messungen aus dem Monitoring auch Analysen von C, P und N der organischen Partikel im Epilimnion nötig. Wie sich in den letzten Jahren bestätigt hat, verändern sich die Verhältnisse zwischen Nährstoffen, Biomasseproduktion und Stöchiometrie im Verlauf der Oligotrophierung [7], weshalb sie bei der Bestimmung der NEP berücksichtigt werden müssen.
Die NEP kann in direkter Weise auch mittels Sedimentfallen bestimmt werden, indem die Menge von absinkendem organischem Material pro Fläche und Zeit unmittelbar unter der produktiven Oberflächenschicht aufgefangen wird. Die sorgfältige Wahl der Expositionstiefe und zeitgerechtes Leeren der Fallen sind wichtig, damit kein Material verpasst wird oder schon während der Exposition mineralisiert. Zudem muss auf unerwünschte Einträge von terrestrischem Material geachtet werden.
Zusammenfassend zeigt sich, dass die aktuell erprobten Methoden zur Bestimmung der PP aufwendig und oft wenig repräsentativ sind. Die NEP kann zudem nicht direkt aus der PP abgeleitet werden, weil keine genauen Angaben über die Respiration im Epilimnion vorhanden sind. Für die Auswirkungen des P-Haushalts auf die Produktion und Respiration und somit für die Sauerstoffzehrung im Hypolimnion ist aber die NEP die Schlüsselgrösse. In einer kürzlich an der Eawag durchgeführten Studie sind daher die seit vielen Jahren in den regulären Monitoring-Programmen gemessenen O2-Konzentrationen näher analysiert worden [8, 9]. Auf diese Weise kann die NEP nicht nur aktuell, sondern auch zurückliegend berechnet werden. Aufbauend auf dem Artikel «Sauerstoffzehrung in Seen» [10] wird die Methode nachfolgend dargestellt. Eine ausführliche Arbeitshilfe dazu ist online verfügbar [1].
Beim antagonistischen Aufbau von organischem Material bei Tageslicht und ständiger Mineralisation (Tag und Nacht) bleibt netto eine Menge organischen Materials übrig, die aus der produktiven Zone aussedimentiert und entweder im Hypolimnion abgebaut oder im Sediment deponiert wird (Fig. 1). Aus O2-Messungen des regulären Monitorings kann indirekt nachverfolgt werden, wie viel C im Hypolimnion abgebaut wurde. Die im Sediment eingelagerte Menge C wird in Sedimentkernen gemessen und spielt je nach Seetyp eine unterschiedlich wichtige Rolle. Im Folgenden werden diese Senken vorgestellt und die daraus abgeleitete Berechnung der NEP erklärt.
Anhand der Zehrung des O2, die in direktem Zusammenhang mit dem Abbau des organischen Materials steht (Fig. 1), kann mit dem stöchiometrischen Verhältnis zwischen O2 und C (von 138 : 106) die Menge an mineralisiertem organischem C berechnet werden. Ein Teil des see-internen C aus der NEP wird bereits beim Absinken in der Wassersäule abgebaut (WCM) und als CO2 freigesetzt. Ein weiterer Teil wird unter O2-Zehrung an der Sedimentoberfläche mineralisiert (SOU). Herrschen im Tiefenwasser und am Sediment anoxische Bedingungen, wird organisches Material vermehrt ins Sediment eingelagert und dort langsam ohne O2 abgebaut. Dabei werden über Jahrzehnte reduzierte Substanzen (RED) wie Methan und Ammonium, aber auch Eisen und Mangan freigesetzt. Sobald diese Substanzen mit O2 in Berührung kommen, werden sie oxidiert und tragen somit – zeitlich verzögert – ebenfalls zur O2-Zehrung bei. RED gibt dort somit nicht die aktuelle Produktion wieder, sondern – als «Sediment-Altlast» – diejenige der Vergangenheit. In flachen Seen mit eutropher Vergangenheit ist der Anteil von RED an der O2-Zehrung auch heute noch hoch [10].
Auch eingeschwemmtes, see-externes organisches Material (Blätter, Äste, Humus etc.) trägt zur O2-Zehrung bei. Dieses ist meist schwerer abbaubar [11], sinkt oft rasch zum Grund, tritt mitunter in grossen Mengen auf und kann besonders in nährstoffarmen Seen wesentlich zu RED beitragen.
Nur ein kleiner Teil des see-internen organischen Materials der NEP wird langfristig im Sediment eingelagert (Netto-Sedimentation, NS). Seen mit grosser terrestrischer Fracht aus Zuflüssen weisen generell hohe NS-Werte auf. Der terrestrische Beitrag an der gesamten organischen NS ist naturgemäss nur schwer abschätzbar. Im Folgenden wird die NS daher nur bei Seen berücksichtigt, die im Vergleich zur PP einen kleinen terrestrischen Input aufweisen und wo der Anteil NEP an der NS klar bestimmt werden kann.
Die Zusammensetzung der für die Bestimmung der NEP berücksichtigten Komponenten der C-Senken hängt vom Seetyp ab (Fig. 2). Hat ein flacher See eine eutrophe Vergangenheit, ist das Sediment mit einer grossen Menge nicht-abgebautem organischem Material belastet, die nicht von der aktuellen Produktion stammt. Deshalb wird der Anteil RED von der AHM abgezogen (Typ 1). Ist in einem tiefen See die terrestrische Fracht hoch und kann die NS nicht zweifelsfrei bestimmt werden, wird auch dieser Anteil von der Berechnung ausgeschlossen (Typ 2). Ist ein See tief, mit wenig terrestrischem organischem Material und ohne Sediment-Altlast, so kann die aktuelle NEP vereinfacht als NEP = AHM + NS ausgedrückt werden (Typ 3) [9]. In der Figur 2 ist das Vorgehen für diese drei Seetypen schematisch dargestellt und in der Arbeitshilfe [1] ausführlich erklärt.
Die aerobe Mineralisation von C folgt modellmässig den stöchiometrischen Verhältnissen:
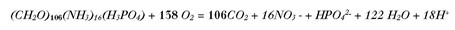
Das C : O2-Verhältnis entspricht demnach 106 : 138. Als Durchschnitt und um den Vergleich mit anderen Seen zu ermöglichen, wird die Dauer der produktiven Phase auf 180 Tage festgelegt. Umgerechnet in eine Produktionsrate pro Flächeneinheit über 180 Tage ergibt dies (hier für Typ 1):
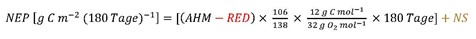
Die Anteile RED (in g O2 m–2 Tag-1) und NS (in g C m–2 [180 Tage]–1) werden je nach Seetyp berücksichtigt. Die AHM kann vereinfacht anhand der Differenz zwischen dem maximalen O2-Gehalt im Hypolimnion nach der Winterzirkulation (etwa Ende März) und dem minimalen O2-Gehalt am Ende der produktiven Saison (Ende Oktober / Anfang November) abgeschätzt werden [12, 13]:
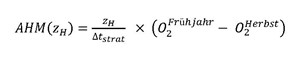
∆tstrat: Schichtungsdauer (in Tagen)
zH: mittlere Hypolimniontiefe (Hypolimnionvolumen dividiert durch Hypolimnionfläche in 15 m Tiefe)
Der von der AHM abzuziehende Anteil RED ist zwar individuell für jeden See, verändert sich aber im Verlauf der Zeit nur langsam [14]. Es ist daher möglich, die in den letzten Jahren durch Porenwasseranalysen ermittelten Werte [8, 9] noch für viele Jahre zu verwenden, denn bis 2050 wird eine Abnahme der RED von nur ~10–15% erwartet. Neben den zur Berechnung der AHM nötigen langen Zeitreihen von O2-Profilen ist es daher ausreichend, einen repräsentativen Wert für RED zu ermitteln.
Auch die durch see-internes Material verursachte NS verändert sich erfahrungsgemäss nur sehr langsam. Generell ist dieser Anteil im Vergleich zur AHM klein. Ist der Wert aus Sedimentproben bekannt, sollte er in die Berechnung miteinbezogen werden. Ist der See stark von terrestrischer Fracht geprägt, so muss diese Komponente ausgeschlossen werden. Werte von RED und NS für verschiedene Seen können in der Arbeitshilfe [1] nachgeschlagen werden. Ebenfalls in der Arbeitshilfe wird eine Typisierung vorgeschlagen, damit nachvollziehbar wird, wann NS und RED berücksichtigt oder vernachlässigt werden sollen.
Die Bestimmung der NEP wird hier am Beispiel des Genfersees (tief, geprägt von see-internem organischem Material) im Detail vorgestellt. Gemäss Figur 2 (Typ 3) wird die NEP als AHM + NS berechnet.
O2-Profile sind im Genfersee seit Ende der 50er-Jahre verfügbar [15]. Anhand der Bathymetriedaten wird zunächst für jedes Messdatum die volumengewichtete, mittlere O2-Konzentration im Hypolimnion (definiert als Tiefe >15 m; Fig. 3a) berechnet. Für jedes Jahr werden das Start- und das Enddatum der produktiven Saison individuell festgelegt. Im Hypolimnion tritt das O2-Maximum zwischen März und Mai auf, das O2-Minimum zwischen September und November. Die O2-Konzentration nimmt während dieser produktiven Schichtungsphase zwischen Start- und Enddatum (Kreise in Fig. 3b) kontinuierlich ab.
Die mittlere Hypolimniontiefe zH beträgt im Genfersee 151 m, die Schichtungsperiode ∆tstrat ist jeweils die Anzahl Tage zwischen den gewählten Start- und Enddaten. Gemäss Gleichung (3) ergibt sich für 1985 (Start: 22.04.1985, Ende: 04.11.1985):
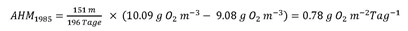
Gleichermassen findet man für 2012 (Start: 09.05.2012, Ende: 23.10.2012) eine AHM2012 von 1.60 g O2 m–2 Tag–1.
Die NS für den Genfersee wurde der Literatur entnommen und beträgt über die produktive Saison ~5,5 g C m–2 (180 Tage)–1 [16]. Mit der Gleichung (2) kann die NEP während der produktiven Phase für jedes Jahr berechnet werden und beträgt für das Beispiel 1985:
Analog dazu beträgt die NEP für 2012 mit 88 g C m–2 (180 Tage)–1 fast doppelt so viel. Wie schon bei der zeitlichen Entwicklung der AHM ist auch für die NEP seit den 1960er-Jahren kein Trend erkennbar (Fig. 4a und 4b). Diese Resultate können mit Sedimentfallenmessungen und den Bilanzen von P und N im Epilimnion überprüft werden. Sedimentdaten von 1993 und 2018 bestätigen, dass sich die NEP in den letzten Jahrzehnten kaum verändert hat und auch die Nährstoffbilanzen der letzten 47 Jahre [9] zeigen eine gute Übereinstimmung mit der NEP in Figur 4b. Der P-Gehalt hat im Gegensatz dazu seit den 1980er-Jahren markant abgenommen (Fig. 4c).
Die einfache Berechnung der NEP nach Gleichung (2) führt bei gewissen Seetypen unter Umständen zu Über- oder Unterschätzung (Fig. 2). Der terrestrische und der aus der PP stammende organische C im See könnten analytisch nur mit sehr grossem Aufwand unterschieden werden. Dies zu vereinfachen, ist Gegenstand aktueller Forschung an der Eawag. Bei flachen Seen ist dies besonders problematisch, da der terrestrische C sowohl die NS als auch RED und SOU erhöht. Abgesehen von terrestrisch geprägten flachen Seen kann die Methode jedoch weitgehend angewendet werden.
Im Vergleich zum Genfersee sind sowohl der Brienzer- als auch der Baldeggersee zwei komplementäre Fälle. Der Baldeggersee ist flach, nährstoffreich, belüftet und hat eine hocheutrophe Vergangenheit. Diese widerspiegelt sich im hohen Anteil von RED, der deshalb nicht zur aktuellen NEP gehört und bei der Anwendung von Gleichung (2) von der AHM abgezogen wird. Die aktuelle NS ist vermutlich noch ähnlich wie zur Zeit der Hocheutrophie und ihr Anteil wird deshalb dazugerechnet [9]. Der Baldeggersee entspricht dem Typ 1 in Figur 2. Der Brienzersee ist tief, nährstoffarm und weist eine geringe PP auf. Er ist jedoch stark geprägt durch grosse Frachten von terrestrischem organischem Material. Damit entspricht er dem in Figur 2 charakterisierten Typ 2. Sedimentfallenmessungen haben gezeigt, dass dieser Eintrag für die gesamte NS und daher auch für RED verantwortlich ist (Fig. 5). Über lange Zeiträume trägt ein Teil dieses externen Materials auch zur SOU bei, wodurch die berechnete NEP etwas überschätzt wird [9].
Die vorgeschlagene Methode erweist sich bei beiden Seentypen als anwendbar: So hat zum einen die PP im Baldeggersee noch nicht messbar abgenommen und zum anderen werden im Brienzersee diejenigen NEP-Anteile, die nicht durch see-interne PP verursacht sind, von der Berechnung ausgeschlossen.
Der Bodensee weist eine sehr ähnliche Entwicklung auf wie der im Detail diskutierte Genfersee. Er ist ebenfalls tief und erhält eine geringe Fracht von terrestrischem organischem C, jedoch sind die P-Konzentrationen geringer als im Genfersee (Fig. 6). Die NEP im Bodensee liegt aktuell bei 56 g C m–2 (180 Tage)–1 [9]. Beide Seen entsprechen dem Typ 3 in Figur 2.
Für die Intensität der Prozesse im Hypolimnion eines Sees ist nicht so sehr die Primärproduktion (PP) von Biomasse wichtig, sondern vor allem jener Anteil, der letztlich aus der produktiven Zone ins Hypolimnion absinkt. Mehr als die schwer messbare PP ist daher die Netto-Ökosystemproduktion (NEP) direkt massgebend für die Sauerstoffzehrung, für die Ablagerung von organischem Kohlenstoff in den Sedimenten und für die Nährstoffkreisläufe (Elimination von Stickstoff durch Denitrifikation, Ablagerung von Phosphor). Anstelle der PP, für welche die Langzeitdaten meist fehlen, kann mit der NEP die trophische Entwicklung eines Sees über längere Zeit zurückverfolgt und daher auch langsame Veränderungen erkannt werden. Die vorgestellte Methode erlaubt es, anhand von Daten der regulären Monitoring-Programme mit wenig Aufwand die NEP eines Sees zu bestimmen. Dabei erhält man einen integrierten Wert über die gesamte produktive Saison ohne feinere zeitliche Auflösung. Obwohl die NEP in manchen Seen etwas unter- oder überschätzt werden kann, erlaubt sie, die zeitliche Entwicklung über Jahre robust zu dokumentieren. Es wird somit ersichtlich, ob die NEP und damit auch die PP längerfristig abnimmt und Oligotrophierung stattfindet, wie viel zur angestrebten «mittleren Produktion» noch fehlt oder ob trotz abnehmendem Nährstoffgehalt keine Abnahme der PP zu erkennen ist. Die Analyse zeigt beispielsweise, dass der Baldeggersee nach wie vor eine maximale PP aufweist, während beim Genfersee erst ein geringer und beim Bodensee ein deutlicher Rückgang der Primärproduktion zu erkennen ist. Die NEP ergänzt und erweitert die Information aus der Sauerstoffzehrung im Hypolimnion (AHM), indem sie die Produktion auf die Basis von Kohlenstoff stellt und dabei die Speicher bzw. Quellen von Sediment und see-externen Einflüssen miteinbezieht. Dadurch werden unterschiedliche Seen miteinander vergleichbar und der Erfolg der Anstrengungen des Gewässerschutzes in den Einzugsgebieten sichtbar. Nährstoffbilanzen ermöglichen einen unabhängigen Plausibilitätscheck der berechneten NEP, jedoch ist die Bestimmung aus den O2-Zehrungsdaten in den meisten Fällen genauer und einfacher. Bei grosser Diskrepanz zwischen den verschiedenen Berechnungsmethoden kann der Grund für die Differenz eine Hilfe für Fehlerabschätzungen sein oder um ungenügende Informationen über Frachten zu erkennen. Die Anleitung zur Berechnung aller in diesem Artikel vorgestellten Methoden sowie der tolerierbaren Werte von NEP, AHM und Phosphoreintrag für einen individuellen See kann der Arbeitshilfe [1] entnommen werden.
[1] Kiefer, I.; Müller, B.; Wüest, A. (2021): Anleitung zur Analyse von Sauerstoffzehrung und Netto-Ökosystemproduktion (2021). Arbeitshilfe zur Ermittlung relevanter Grössen der Trophie von Seen, Eawag und EPFL
[2] Marra, J. (2009): Net and gross productivity: weighing in with 14C. Aquatic Microbial Ecology 56, 123–131
[3] Slawyk, G.; Collos, Y.; Auclair, J-C. (1977): The use of the 13C and 15N isotopes for the simultaneous measurement of carbon and nitrogen turnover rates in marine phytoplankton, Limnol. Oceanogr. 22(5), 925–932
[4] Brothers, S. et al. (2017): Convective mixing and high littoral production established systematic errors in the diel oxygen curves of a shallow, eutrophic lake. Limnol. Oceanogr.: Methods 15, 429–435
[5] Odermatt, D. et al. (2012): Review of constituent retrieval in optically deep and complex waters from satellite imagery. Remote Sens Environ. 118, 116–126
[6] Soomets, T. et al. (2019): Spatial and temporal changes of primary production in a deep peri-alpine lake. Inland Waters 9(1), 49–60
[7] Müller, B. et al. (2021): Increasing carbon-to-phosphorus ratio (C:P) from seston as a prime indicator for the initiation of lake reoligotrophication, im Druk, Environ. Sci. Technol.
[8] Steinsberger, T. et al. (2020): Hypolimnetic oxygen depletion rates in deep lakes: Effects of trophic state and organic matter accumulation. Limnol. Oceanogr. 65(12), 2020, 3128–3138
[9] Steinsberger, T.; Wüest, A.; Müller, B. (2021): Net ecosystem production of lakes estimated from hypolimnetic organic carbon sinks, eingereicht bei Water Resources Research
[10] Kiefer, I. et al. (2020): Sauerstoffzehrung in Seen, Aqua & Gas N°7/8, 62–70
[11] Meyers, P.A.; Eadie, B.J. (1993): Sources, degradation and recycling of organic matter associated with sinking particles in Lake Michigan. Org. Geochem. 20(1), 47–56
[12] Matzinger, A. et al. (2010): Hypolimnetic oxygen consumption by sediment-based reduced substances in former eutrophic lakes. Limnol. Oceanogr. 55 (5), 2073–2084
[13] Müller, B. et al. (2012): Hypolimnetic oxygen depletion in eutrophic lakes. Environ. Sci. Technol. 46(18), 9964–9971
[14] Steinsberger, T. et al. (2019): Modeling sediment oxygen demand in a highly productive lake under various trophic scenarios. PLoS ONE 14(10): e0222318
[15] Rimet, F. et al. (2020): The Observatory on LAkes (OLA) database: Sixty years of environmental data accessible to the public. Journal of Limnology 79(2), 164–178
[16] Steinsberger, T. et al. (2017): Organic carbon mass accumulation rate regulates the flux of reduced substances from the sediments of deep lakes. Biogeosciences 14(13), 3275–3285
[17] Rapin, F.; Gerdeaux, D. (2013): La protection du Léman, priorité à la lutte contre l’eutrophisation. Arch.Sci. 66, 103–116
Dieser Artikel wurde im Rahmen des vom BAFU finanzierten Projekts «Primärproduktion der Seen unter Oligotrophierung: Verfahren zur Erhebung des Produktionsstatus basierend auf Routineuntersuchungen und öffentlich zugänglichen Daten (Trophie-Status)» erstellt. Forschungsarbeiten wurden vom Schweizerischen Nationalfonds (Projekte: 200021_146652, 200020_165517 und 200021_146234) unterstützt. Wir bedanken uns zudem herzlich bei den kantonalen Umweltämtern (AfU Freiburg; Amt für Landwirtschaft und Umwelt Obwalden; Amt für Umwelt Nidwalden; Amt für Umweltschutz Schwyz; Amt für Wasser und Energie St. Gallen; Amt für Umwelt Zug; AWA Bern; AWEL Zürich; Departement Bau, Verkehr und Umwelt Aargau; DGE Vaud; Service de l’énergie et de l’environnement Neuchâtel; uwe Luzern), bei der IGKB Langenargen, der CIPAIS, der CIPEL (mit Informationssystem Observatory of alpine LAkes (OLA), © SOERE OLA-IS, AnaEE-France, INRA in Thonon-les-Bains, CIPL) sowie dem BAFU für die jahrzehntelange Durchführung unzähliger Messungen in den Schweizer Seen und die daraus zur Verfügung gestellten wertvollen Datensätze.
«AQUA & GAS» gibt es auch als E-Paper. Abonnenten, SVGW- und/oder VSA-Mitglieder haben Zugang zu allen Ausgaben von A&G.
Den «Wasserspiegel» gibt es auch als E-Paper. Im SVGW-Shop sind sämtliche bisher erschienenen Ausgaben frei zugänglich.




Kommentare (0)