Plateforme pour l’eau, le gaz et la chaleur
")
, * sites à bassin versant agricole (AGR). 0+: truites de l’année, 1+: truites d’un an. N.d.: le sexe n’a pas pu être déterminé. T°C: température, O2 : oxygène dissous, Cond.: conductivité.")
![Fig. 1 Préparation des échantillons et mesure des biomarqueurs par qPCR microfluidique.
1. L’ARN total est extrait des tissus de cerveau et de foie des truites de rivière.
2. L’ARN messager est converti en ADN complémentaire.
3. Les échantillons et les paires d’amorces spécifiques aux 96 biomarqueurs sont chargés sur une plaque microfluidique. Les réactions de qPCR sont mélangées dans des nanochambres au centre de cette plaque. La fluorescence émise par la réaction est enregistrée en temps réel par l’instrument Biomark HD (Fluidigm). Pour chaque échantillon, une valeur de Ct est obtenue, qui équivaut au nombre de cycles nécessaires pour détecter l’amplification du biomarqueur et permet de quantifier son expression. (Adapté de [32])](/media/1234463/fa_voisin_fig-1.png?width=760&height=440&bgcolor=e8e8e8 "Fig. 1 Préparation des échantillons et mesure des biomarqueurs par qPCR microfluidique.
1. L’ARN total est extrait des tissus de cerveau et de foie des truites de rivière.
2. L’ARN messager est converti en ADN complémentaire.
3. Les échantillons et les paires d’amorces spécifiques aux 96 biomarqueurs sont chargés sur une plaque microfluidique. Les réactions de qPCR sont mélangées dans des nanochambres au centre de cette plaque. La fluorescence émise par la réaction est enregistrée en temps réel par l’instrument Biomark HD (Fluidigm). Pour chaque échantillon, une valeur de Ct est obtenue, qui équivaut au nombre de cycles nécessaires pour détecter l’amplification du biomarqueur et permet de quantifier son expression. (Adapté de [32])")
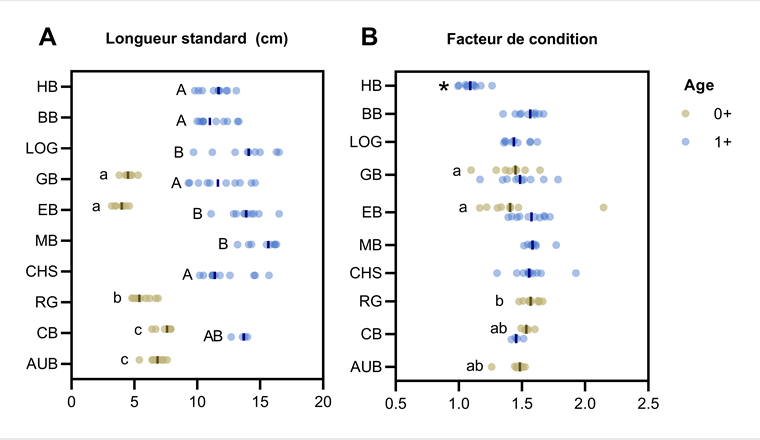
 et facteur de condition (B). Les valeurs individuelles et la médiane sont représentées pour chaque site. Les sites sont ordonnés par date d’échantillonnage. Pour une même catégorie d’âge, des lettres différentes indiquent une différence significative entre sites; * indique une différence significative par rapport à tous les autres sites (p < 0,05).")
Les méthodes basées sur les effets biologiques (effect-based methods) constituent un outil prometteur permettant de faire le lien entre l’exposition aux mélanges complexes de micropolluants et les conséquences pour les organismes aquatiques [1-3]. Ces méthodes regroupent entre autres les bioessais, tests standardisés en laboratoire réalisés à partir de prélèvements d’eau, et les biomarqueurs.
Un biomarqueur est une réponse biologique quantifiable suite à un stress environnemental [4]. Ces réponses peuvent être mesurées à différents niveaux: expression des gènes, activité enzymatique, paramètres physiologiques, sur les organismes que ce soit en laboratoire ou sur les organismes prélevés sur le terrain [5, 6] et représenter de nombreux processus biologiques (par ex. métabolisme, système immunitaire, neurotoxicité). Les biomarqueurs analysés chez les poissons sauvages reflètent directement leur état de santé et peuvent indiquer la nature des facteurs de stress auxquels les animaux ont été et/ou sont exposés [3, 7].
Les changements dans l’expression de gènes font partie des premières réponses cellulaires suivant un stress tel que l’exposition à des micropolluants [5] et peuvent servir de biomarqueurs moléculaires permettant de détecter les effets sublétaux de façon sensible et précoce.
La méthode de référence pour mesurer l’expression des gènes est la réaction en chaîne par polymérase quantitative (qPCR) [8]. Cette méthode peut être coûteuse en termes de matériel et de temps si l’on souhaite mesurer un large panel de biomarqueurs et de nombreux échantillons. Un large set de biomarqueurs est cependant nécessaire pour détecter au mieux les réponses biologiques induites par les stress environnementaux [9]. Ces dernières années, des méthodes à haut débit ont été largement développées. Par exemple, le système de qPCR microfluidique BioMark HD (Standard BioTools/Fluidigm, San Francisco, USA) permet de mesurer 96 biomarqueurs sur 96 échantillons en une seule analyse, soit 9216 mesures, contre 384 mesures en qPCR classique. De plus, cette technique permet de réduire la quantité de réactifs et d’échantillon nécessaire. Ceci est avantageux lorsque la quantité de matériel biologique est limitée, notamment lorsque l’on étudie les poissons juvéniles.
Les premiers stades de vie sont particulièrement sensibles aux stress environnementaux. En Suisse, les truites de rivière (Salmo trutta) fraient de novembre à janvier, et les premiers stades de vie de la truite sont donc potentiellement présents dans l’eau au printemps et au début de l’été lorsque le risque de contamination par les pesticides est le plus élevé pour les poissons [10–12].
Dans ce contexte, notre étude vise à:
Le foie et le cerveau ont été choisis, le foie étant l’organe principal du métabolisme, notamment de biotransformation des substances toxiques. Le cerveau a été analysé afin de détecter les potentiels effets neurotoxiques dus à la présence de pesticides.
Dans cet article, nous présentons les résultats de cette étude pilote. En coordination avec une batterie de bioessais réalisés à partir d’échantillons d’eau et de sédiments prélevés à ces mêmes sites dans le cadre de deux autres projets menés au Centre Ecotox [13-15], ainsi que d’une analyse chimique approfondie, nous cherchons à établir une image la plus complète de la qualité des eaux et faire un pas vers l’utilisation des biomarqueurs en biosurveillance.
Un total de dix sites répartis sur cinq cantons, dont cinq sites à bassin versant agricole (AGR) et cinq sites à bassin versant exploité de manière extensive (EXT), ont été sélectionnés en coordination avec deux autres projets menés au Centre Ecotox [14, 15]. La description complète des sites est disponible dans un article commun dans ce même numéro (voir p. 18, [13]). Des truites de rivière (S. trutta) juvéniles ont été échantillonnées par pêche électrique avec l’aide des gardes-pêche cantonaux entre mai et fin juillet 2021, période correspondant à une forte application de pesticides [12]. Le tableau 1 reprend les sites, la date de pêche, le nombre de truites échantillonnées ainsi que les caractéristiques physicochimiques de l’eau. En raison de la sensibilité des premiers stades de vie, nous avons cherché à échantillonner dix truites de l’année (0+) par site. À cinq sites (HB, LOG, BB, MB et CHS), les 0+ n’ont pas pu être observées, dès lors des truites d’un an (1+) ont été prélevées. À EB, GB et CB, les deux catégories d’âge ont été échantillonnées. À AUB et RG, seules les truites 0+ ont été prélevées.
Les poissons ont été euthanasiés sur place par une surdose de méthanesulfonate de tricaïne (MS222) suivie d’une coupe des branchies. La longueur standard et le poids ont été mesurés, puis le foie et le cerveau ont été disséqués et immédiatement congelés dans de la glace sèche jusqu’au stockage en laboratoire à -80 °C. Le facteur de condition de Fulton (FC) a été calculé comme le rapport entre le poids et la longueur du poisson. Il donne une indication générale de l’état de santé des poissons, notamment de leur état de nutrition et de maturité sexuelle.
À chaque site, des échantillons d’eau composites de 14 jours précédant la capture des poissons ont été prélevés pour l’analyse de bioessais et l’analyse chimique des micropolluants organiques, des paramètres abiotiques et des métaux [13, 14]. De plus, des bioessais et une analyse chimique ont été réalisés à partir de sédiments prélevés à ces mêmes sites [15].
Un ensemble de 96 gènes biomarqueurs a été sélectionné avec objectif de détecter les effets sublétaux de micropolluants, en particulier des pesticides, et d’autres facteurs de stress (par ex. la température et l’infection par des agents pathogènes). Nous avons inclus des biomarqueurs développés en collaboration avec l’Eawag et aQuaTox Solutions GmbH lors de projets précédents [16, 17] et d’une étude en laboratoire avec la truite de rivière [18]. Ce set a été élargi en comparant les informations de la litérature scientifique [19–23] et de bases de données
(ctdbase.org et aopwiki.org) [24]. Les biomarqueurs sélectionnés couvrent de nombreux processus biologiques:
Un micropolluant qui se retrouve dans l’organisme peut être détoxifié par le métabolisme de biotransformation (neuf biomarqueurs mesurés). Ce processus facilite l’élimination des substances chimiques en augmentant leur solubilité. Par exemple, le récepteur d’aryl hydrocarbone (codé par le gène ahr), lorsqu’il détecte certaines substances chimiques dans la cellule, active l’expression des gènes de biotransformation tels que cyp1a. La biotransformation joue également un rôle dans la toxicité de certains composés chimiques (ex. insecticides organophosphates, dioxines et hydrocarbures aromatiques polycycliques HAP) par la formation de métabolites plus toxiques (bioactivation) [25].
Le stress oxydant (six biomarqueurs) se produit lorsqu’il y a une surproduction d’espèces réactives de l’oxygène, des molécules pouvant endommager les cellules et les protéines. Le stress oxydant est un effet secondaire fréquent du métabolisme de biotransformation et un mécanisme important de toxicité des micropolluants [26, 27], dont les insecticides pyréthrinoïdes [21, 28]. Le stress oxydant peut avoir des conséquences directes ou indirectes sur plusieurs processus cellulaires (cinq biomarqueurs). Par exemple, il peut activer l’apoptose (mort cellulaire), qui élimine les cellules endommagées.
De nombreux micropolluants peuvent induire l'inflammation et impacter d'autres acteurs du système immunitaire (neuf biomarqueurs), notamment suite au stress oxydant.
La mort cellulaire et l’inflammation dans le système nerveux sont des mécanismes courants de neurotoxicité (quinze biomarqueurs) [21]. Celle-ci elle peut aussi résulter de l’interaction directe des micropolluants avec des acteurs du système nerveux. C’est le cas de l’inhibition de l’enzyme acétylcholinestérase (AChE, codée par le gène ache) par les insecticides organophosphorés et carbamates, ou du blocage des canaux sodium par les pyréthrinoïdes, entrainant la paralysie et la mort de l’organisme.
De nombreuses substances chimiques qui se retrouvent dans l’environnement sont capables d’interférer avec le métabolisme des hormones ou encore de se lier à leurs récepteurs [29]. On parle de perturbation endocrinienne (onze biomarqueurs).
Certains pesticides sont susceptibles d’alterer le métabolisme du cholestérol et des stéroides (quatre biomarqueurs) [19, 30]. Notamment, les fongicides azolés inhibent l’enzyme stérol 14α-déméthylase, essentielle au fonctionnement des membranes chez les champignons [31]. Cette enzyme, codée par le gène cyp51, est également présente chez les vertébrés.
Afin de compléter le diagnostic de l’état de santé des organismes, nous avons inclus six biomarqueurs du métabolisme des lipides et quatre du métabolisme énergétique qui ont des implications importantes pour la croissance et la reproduction des organismes.
Enfin, nous avons inclus des biomarqueurs permettant de détecter une potentielle exposition aux métaux (quatre biomarqueurs) et une réponse générale au stress et au stress thermique (cinq biomarqueurs).
Après extraction de l’ARN du foie et du cerveau, les échantillons ont été normalisés à la même concentration, puis l’ADN complémentaire a été synthétisé à partir de l’ARN messager (fig. 1). Les échantillons des 117 truites en duplicat technique ainsi que les 96 paires d’amorces (courtes séquences d’ADN permettant la mesure des 96 biomarqueurs) ont été chargés sur un total de trois plaques microfluidiques pour chaque organe. L’expression des biomarqueurs a été calculée à partir des valeurs de Ct (cycle threshold ou cycle seuil) selon la méthode décrite dans Taylor et al. [8]. Pour chaque biomarqueur, nous avons calculé le log2 de l’expression relative à la moyenne de tous les échantillons, normalisée avec trois gènes de référence (arp, ubi, rpl7). Pour cette première étude de terrain, nous avons mesuré le même set de biomarqueurs dans le foie et le cerveau. Après exclusion des gènes peu ou pas exprimés dans chaque organe, nous obtenons les résultats de 79 biomarqueurs dans le foie et de 89 biomarqueurs dans le cerveau.
L’analyse statistique a été réalisée à l’aide du logiciel GraphPad Prism (v. 9.4.1) et porte sur les aspects suivants:
Une différence significative de longueur standard des truites a été observée entre plusieurs sites dans les deux catégories d’âge (fig. 2A). Ces différences peuvent être dues à la variabilité entre populations et à la croissance des truites entre les échantillonnages (les sites AUB, CB, RG ayant été échantillonnés fin juillet par exemple). Représentant le rapport entre le poids et la taille des truites, le facteur de condition (fig. 2B) était significativement plus petit à HB (1+) par rapport à tous les autres sites. Un facteur de condition plus faible chez ces truites pourrait être un premier indicateur d’un état de santé moins bon et/ou d’un état de maturité moins avancé.
L’expression des gènes étant à la base de presque tout processus biologique, y compris le développement et la reproduction, il est attendu que l’expression de certains biomarqueurs soit influencée par l’âge et le sexe des individus. Parmi les biomarqueurs mesurés, neuf biomarqueurs dans le foie et 34 dans le cerveau étaient significativement corrélés à la longueur standard, et sont donc susceptibles de varier selon le développement et l’âge des truites. De plus, seize biomarqueurs dans le foie et neuf biomarqueurs dans le cerveau étaient différentiellement exprimés selon le sexe, dont plusieurs gènes impliqués dans la reproduction. Compte tenu de ces résultats, avons examiné les différences entre sites en considérant chaque catégorie d’âge séparément et en tenant compte de la taille et du sexe lors de l’interprétation des réponses.
Au vu des concentrations parfois élevées de pesticides et de métaux [14], les sites à bassin versant exploité de manière extensive ne peuvent pas servir de référence pour l’expression des biomarqueurs. Nous avons dès lors cherché à mettre en évidence les effets biologiques en comparant les valeurs de biomarqueurs entre tous les sites pour chaque catégorie d’âge. Nous considérons un effet lorsque l’expression du biomarqueur est significativement sur- ou sous-exprimé par rapport à la majorité des autres sites. Les biomarqueurs dont l’expression varie fortement entre de nombreux sites n’ont pas été considérés, car en l’absence de valeurs de référence, il est impossible de savoir si les résultats indiquent un état normal ou stressé. Le tableau 2 donne une vue d’ensemble des effets biologiques mis en évidence.
Les biomarqueurs de biotransformation cyp3a et abcb1 (foie) étaient significativement sur-exprimés chez les truites de CHS par rapport aux autres sites, tandis que cyp1a (cerveau) était sur-exprimé à CB et sous-exprimé à HB. Cyp1a suit la même tendance dans le foie (non significatif).
Le biomarqueur nrf2, orchestrateur des défenses antioxydantes, était significativement sur-exprimé chez les truites de CHS et CB (foie, 1+). Le gène codant pour la glutathion réductase (gsr, foie), responsable de restaurer les réserves en glutathion nécessaire à la biotransformation et aux défenses antioxydantes, était sur-exprimé à EB (1+) et le gène codant pour la catalase (cat, foie) était sous-exprimé à HB.
Les gènes impliqués dans le système nerveux bdnf et calm1a étaient respectivement sur- et sous-exprimés à CB (cerveau, 1+). La réponse chez les 0+ suit la même tendance, bien que statistiquement non significative. Le gène codant pour le récepteur GABAA (gabra1), cible moléculaire de certains pesticides tels que le fipronil [33], était significativement sous-exprimé chez les truites 1+ à MB, CB et CHS.
Parmi les gènes impliqués dans le système immunitaire, b2m, socs3 et il6 (foie) étaient sur-exprimés à CB (0+ et 1+), et socs3 à HB et CHS.
Des effets sur le système endocrinien ont été observés chez les truites de HB avec une sous-expression des gènes codants pour l’aromatase, le récepteur aux androgènes (cyp19a1b et ar, cerveau) et les récepteurs aux glucocorticoïdes et minéralocorticoïdes (gr et mr, foie). Un biomarqueur du sytème hormonal thyroïdien (dio2, foie) était sous-exprimé à HB, CHS et AUB.
Plusieurs biomarqueurs du métabolisme du cholestérol et des stéroïdes (cyp51, hsd17b7 et srebf2, foie) étaient fortement sous-exprimés à CB (0+ et 1+) par rapport à tous les autres sites. Plusieurs gènes du métabolisme des lipides étaient sous-exprimés chez les truites 1+ de HB (fasn, acaa1, srebf1), MB et CB (fabp1, fabp3) et sur-exprimés à BB (fabp1, fabp3) et EB (fasn, fabp3). Un biomarqueur du métabolisme énergétique, g6pd, était fortement sur-exprimé à EB (1+).
Le gène codant pour la métallothionéine (mt2, foie) était sur-exprimé à MB, suggérant une exposition aux métaux.
Le gène hsp70 signale le niveau de dommages cellulaires infligé par un ou plusieurs facteurs de stress et est particulièrement sensible au stress thermique [34]. Hsp70 (foie et cerveau) était fortement sur-exprimé à CB, où la température lors de l’échantillonnage était la plus élevée (16,6 °C), suivi de CHS (15,8 °C). Cet effet n’a cependant pas été observé à RG (15,4 °C). Nous pourrions émettre l’hypothèse que des stress multiples soient responsables de la sur-expression de hsp70 à CB et CHS, mais pas à RG.
Selon l’analyse des biomarqueurs, les truites échantillonnées aux sites CB, CHS, HB, EB et MB semblent les plus impactées. La majorité des effets ont été attribués aux sites à bassin versant agricole, mais plusieurs effets ont été observés aux sites à bassin versant exploité de manière extensive. Pour les sites où les deux catégories d’âge ont été échantillonnées, la plupart des effets ont été identifiés chez les truites 1+ (EB et CB; tab. 2). Alors que nous cherchions initialement à prélever les truites 0+, nos résultats indiquent que l’analyse de biomarqueurs sur les truites 1+ est également pertinente.
Dans ce paragraphe, nous comparons les résultats de l’analyse chimique et des bioessais [13–15] afin d’identifier les complémentarités entre les différentes approches et de chercher à expliquer les effets observés sur les biomarqueurs. Il est important de garder à l’esprit les disparités entre ces approches: entre autres, les espèces considérées et leur sensibilité, à la détection ou non de stress multiples et de facteurs confondants, la sensibilité de la méthode, la dépendance au moment d’échantillonnage et au traitement des échantillons, etc.
Suite à l’analyse chimique des échantillons d’eau, le risque aigu et chronique a été analysé pour les plantes, les invertébrés et les vertébrés (poissons et amphibiens) en comparant les concentrations mesurées aux critères de qualité environnementale [14]. Seul le site HB a été identifié comme présentant un risque chronique élevé pour les vertébrés, attribuable principalement aux fortes concentrations de deltaméthrine. C’est également à HB que nous avons observé un facteur de condition des truites 1+ significativement plus faible par rapport aux autres sites. Le sexe de huit des dix truites 1+ n’a pas pu être déterminé, indiquant un possible retard de maturité par rapport aux autres sites. Les biomarqueurs indiquent une possible perturbation endocrinienne (par ex. sous-expression de gènes impliqués dans la reproduction) et une réduction du métabolisme des lipides. Dans le cas de HB, l’analyse du risque chimique et les résultats biomarqueurs semblent concorder. Néanmoins, les biomarqueurs indiquent des effets sublétaux à plusieurs autres sites (MB, CB, CHS, EB) où aucun risque chimique n’a été identifié pour les vertébrés. Concernant les invertébrés, un risque chronique, majoritairement attribuable aux insecticides, a été identifié à MB, CB, et EB. Il est possible que la contamination à ces sites affecte les truites en provoquant des effets directs sublétaux, tels que détectés par les biomarqueurs, et/ou de façon indirecte en impactant les invertébrés, faisant partie de la source de nourriture des truites [22].
Les métaux et autres paramètres abiotiques mesurés dans l’eau (échantillons ponctuels et/ou composites) ont quant à eux montré des dépassements des exigences de l’Ordonnance sur la protection des eaux pour le cuivre (MB, CB, EB, RG), le zinc (RG), le nitrate (EB), et le carbone dissous (CB). Dans les sédiments, le cuivre dépassait le critère de qualité pour le site MB [15]. Ces résultats concordent avec la sur-expression de métallothionéine (mt2) à MB. L’expression de mt2 était également élevée à CB et RG (non significatif).
Certains des effets mesurés par les biomarqueurs peuvent être comparés aux résultats des bioessais [14], tels que les bioessais in vitro mesurant l’activation ou l’inhibition de récepteurs présents chez les vertébrés, les tests in vivo sur embryons [35] et in vitro sur lignée cellulaire de poisson [36]. Le bioessai Nrf2-CALUX® a par exemple identifié un stress oxydant à MB, CB, CHS et RG. En comparaison, nous avons observé une sur-expression de nrf2 chez les truites de CB et CHS. Le bioessai PXR-CALUX® a répondu à de nombreux sites, tandis que l’expression du gène biomarqueur pxr n’a pas montré de réponse claire. Au niveau des effets de perturbation endocrinienne, les bioessais ER- et Anti-AR-CALUX® n’ont pas montré d’effet, alors que les biomarqueurs ont indiqué plusieurs effets sur le système endocrinien, principalement à HB. La toxicité aigüe et chronique des échantillons d’eau a été évaluée par biotest in vivo avec des embryons de poisson zèbre [35]. Notamment, des effets sub-létaux importants ont été attribués aux sites HB, LOG, MB (EXT), BB, CB, EB et RG (AGR). De plus, GB, LOG et tous les sites AGR ont montré une toxicité sur une lignée cellulaire de branchies de truite arc-en-ciel.
Les résultats de l’analyse du risque chimique, des bioessais et des biomarqueurs ne concordent pas dans tous les cas. Combiner ces approches complémentaires permet dès lors d’augmenter les chances de détecter les risques écotoxicologiques et de préciser les effets biologiques. Selon l’analyse chimique, seul le site HB serait considéré comme à risque pour les vertébrés. Selon les résultats des biomarqueurs et de certains bioessais [14], il serait judicieux d’investiguer d’autres sites tels que MB, CB, CHS et EB. Plusieurs autres études récentes ont montré le potentiel de combiner l’analyse chimique et la mesure de biomarqueurs en biosurveillance [30, 37].
Les biomarqueurs mesurés chez les organismes résidents apportent une plus-value importante aux bioessais et à l’approche conventionnelle basée sur l’analyse du risque chimique, car ils permettent de couvrir une très large gamme de processus biologiques au sein d’organismes exposés de façon continue à l’ensemble des conditions environnementales, intégrant les mélanges de contaminants biodisponibles et leurs interactions, les paramètres abiotiques, les effets de la température et les effets indirects (ex. impacts sur la chaîne trophique). Bien que l’application des biomarqueurs en biosurveillance de routine est soutenue depuis de nombreuses années par la communauté scientifique [1, 9, 38–40], elle en est encore à ses débuts. Dans cette étude, nous avons fait un pas dans cette direction en développant un outil permettant la mesure rapide et précise d’une centaine de gènes biomarqueurs chez la truite de rivière, que nous avons appliqué avec succès à des échantillons de terrain. Nos résultats démontrent le potentiel de cet outil en contribuant de manière déterminante à l’identification des sites problématiques (hot spots) et des facteurs de stress pertinents. Notre étude a également permis de mettre en évidence des facteurs confondants (âge, sexe) qui doivent être pris en compte lors de l’analyse des données d’expression génique. L’absence de valeurs de référence freine actuellement l’interprétation des résultats: il faudrait en effet pouvoir dissocier la variation naturelle, due au stade de vie, aux facteurs génétiques et aux facteurs abiotiques (saison, physicochimie), des valeurs d’expression résultant d’un stress [41]. Déterminer ces valeurs de référence, comme l’a fait une étude en France chez le gammare (Gammarus fossarum) et la moule zèbrée (Dreissena polymorpha) [41], nécessiterait la mesure des biomarqueurs ainsi que des facteurs confondants sur un grand nombre de sites. Les travaux futurs devraient viser à déterminer ces valeurs afin d’exploiter pleinement le potentiel des biomarqueurs. L’outil développé dans notre étude permettrait d’obtenir ce large set de données.
[1] Milinkovitch, T. et al. (2019): Biomarkers as tools for monitoring within the Water Framework Directive context: concept, opinions and advancement of expertise. Environ. Sci. Pollut. Res. 26: 32759–32763
[2] Brack, W. et al. (2019): Effect-based methods are key. The European Collaborative Project SOLUTIONS recommends integrating effect-based methods for diagnosis and monitoring of water quality. Environ. Sci. Eur. 31: 4–9
[3] Connon, R.E. et al. (2012): Effect-based tools for monitoring and predicting the ecotoxicological effects of chemicals in the aquatic environment. Sensors (Switzerland) 12: 12741–12771
[4] Beiras, R. (2018): Biological Tools for Monitoring. Mar. Pollut. 265–291, doi:10.1016/b978-0-12-813736-9.00016-7
[5] Schirmer, K. et al. (2010): Transcriptomics in ecotoxicology. Anal. Bioanal. Chem. 397: 917–923
[6] Hasenbein, M. et al. (2014): Transcriptomic profiling permits the identification of pollutant sources and effects in ambient water samples. Sci. Total Environ. 468–469: 688–698
[7] Frank, D. F. et al. (2018): Bifenthrin causes transcriptomic alterations in mTOR and ryanodine receptor-dependent signaling and delayed hyperactivity in developing zebrafish (Danio rerio). Aquat. Toxicol. doi:10.1016/j.aquatox.2018.04.003
[8] Taylor, S. C. et al. (2019): The Ultimate qPCR Experiment: Producing Publication Quality, Reproducible Data the First Time. Trends in Biotechnology 37: 761–774
[9] Lam, P.K.S.; Gray, J.S. (2003): The use of biomarkers in environmental monitoring programmes. Mar. Pollut. Bull. 46: 182–186
[10] Schneeweiss, A. et al. (2019): Ökotoxikologische Risiken von psm für Fische. Aqua & Gas 11/2019: 74–80
[11] Werner, I.et al. (2021): Environmental Risk of Pesticides for Fish in Small- and Medium-Sized Streams of Switzerland. Toxics 9: 79
[12] Spycher, S. et al. (2019): Anhaltend hohe PSM-Belastung in Bächen. Aqua & Gas 4/2019: 14–25
[13] Kienle, C. et al. (2023): Ökotoxikologische Biotests und Biomarker. Aqua & Gas 4/2023: 18–22
[14] Kienle, C. et al. (2023): Beurteilung der Wasserqualität mittels Biotestbatterie. Aqua & Gas 4/2023: 24–33
[15] Casado-Martinez, M. C. et al. (2023): Évaluation de la qualité des sédiments - Projet pilote d'application d’une batterie de bioessais à l'échelle nationale. Aqua & Gas 4/2023: 34–41
[16] Fischer, S.et al. (2017): Wirkungsorientierte Gewässerüberwachung: Biomonitoring mit Forellen. Studie im Auftrag des Bundesamtes für Umwelt 117
[17] Zöllig, H. et al. (2017): PAK-Stufe ARA Herisau. Aqua & Gas 1/2017: 14–23
[18] Rehberger, K. et al. (2021): Schädigen Pflanzenschutzmittel Fische? Aqua & Gas 7/8-2021: 60–69
[19] Uren Webster, T.M. et al. (2015): The Herbicide Linuron Inhibits Cholesterol Biosynthesis and Induces Cellular Stress Responses in Brown Trout. Environ. Sci. Technol. 49: 3110–3118
[20] Akbarzadeh, A. et al. (2018): Developing specific molecular biomarkers for thermal stress in salmonids. BMC Genomics 19: 1–28
[21] Mohammadi, H. et al. (2019): Pyrethroid exposure and neurotoxicity: A mechanistic approach. Arh. Hig. Rada Toksikol. 70: 74–89
[22] Schneeweiss, A.; Werner, I. (2019): Ökotoxikologische Wirkungen von PSM auf Fische. Aqua & Gas 11/2019: 82–91
[23] Ewald, J.D. et al. (2020): EcoToxModules: Custom Gene Sets to Organize and Analyze Toxicogenomics Data from Ecological Species. Environ. Sci. Technol. 54: 4376–4387
[24] Davis, A.P. et al. (2021): Comparative Toxicogenomics Database (CTD): Update 2021. Nucleic Acids Res. 49: D1138–D1143
[25] Schlenk, D. (2005): Chapter 6 Pesticide biotransformation in fish. Biochem. Mol. Biol. Fishes 6: 171–190
[26] Slaninová, A. et al. (2009): A review: Oxidative stress in fish induced by pesticides. Neuroendocrinol. Lett. 30: 2–12
[27] Silvestre, F. (2020): Signaling pathways of oxidative stress in aquatic organisms exposed to xenobiotics. J. Exp. Zool. Part A Ecol. Integr. Physiol. 333: 436–448
[28] Awoyemi, O.M. et al. (2019): Behavioral, molecular and physiological responses of embryo-larval zebrafish exposed to types I and II pyrethroids. Chemosphere 219: 526–537
[29] Brander, S.M. et al. (2016): Pyrethroid pesticides as endocrine disruptors: Molecular mechanisms in vertebrates with a focus on fishes. Environ. Sci. Technol. 50: 8977–8992.
[30] Schmitz, M. et al. (2022): Demonstration of an aggregated biomarker response approach to assess the impact of point and diffuse contaminant sources in feral fish in a small river case study. Sci. Total Environ. 804: 150020
[31] Rösch, A. et al. (2017): Mechanistic Understanding of the Synergistic Potential of Azole Fungicides in the Aquatic Invertebrate Gammarus pulex. Environ. Sci. Technol. 51: 12784–12795
[32] BioRender.com. (2022): Measurement of mRNA Expression Knockdown Using siRNA Through RT-qPCR. https://app.biorender.com/biorender-templates
[33] Stehr, C.M. et al. (2006): The developmental neurotoxicity of fipronil: Notochord degeneration and locomotor defects in zebrafish embryos and larvae. Toxicol. Sci. 92: 270–278
[34] Werner, I. et al. (2005): Heat Shock Proteins in Juvenile Steelhead Reflect Thermal Conditions in the Navarro River Watershed, California. Trans. Am. Fish. Soc. 134:399–410
[35] OECD (2013): Test No. 236: Fish Embryo Acute Toxicity (FET) Test. OECD Publishing, Paris
[36] OECD (2021): Test Guideline No . 249 Fish Cell Line Acute Toxicity: The RTgill-W1 cell line assay. In: OECD guideline for testing of chemicals, OECD Publishing, Paris, doi:10.1787/c66d5190-en
[37] Catteau, A. et al. (2021): Water quality of the Meuse watershed: Assessment using a multi-biomarker approach with caged three-spined stickleback (Gasterosteus aculeatus L.). Ecotoxicol. Environ. Saf. 208: 111407
[38] Sanchez, W.; Porcher, J. (2009): Utilisation des biomarqueurs pour la caractérisation de l’état écotoxicologique des masses d’eau. Tech. Sci. Méthodes, ASTEE/EDP Sci. 5: 29–38
[39] Brack, W. et al. (2017): Towards the review of the European Union Water Framework management of chemical contamination in European surface water resources. Sci. Total Environ. 576: 720–737
[40] Basu, N. et al. (2019): EcoToxChip: A next-generation toxicogenomics tool for chemical prioritization and environmental management. Environ. Toxicol. Chem. 38: 279–288
[41] Leprêtre, M. et al. (2022): Determination of biomarkers threshold values and illustration of their use for the diagnostic in large-scale freshwater biomonitoring surveys. Environ. Sci. Eur. 34: 115
En plus des personnes déjà listées dans [13], les auteurs souhaitent remercier les personnes suivantes pour leurs précieuses contributions: Andreas Scheidegger (Eawag), Bänz Lundsgaard-Hansen (OFEV), Stephan Fischer (aQuaTox-Solutions GmbH), Kristin Schirmer (Eawag), Urs Gsell (Forstbetrieb Muhen-Hirschthal-Holziken) et les gardes-pêche Roman Niedermann, Stefan Eglauf (Ct. Thurgovie), Thomas Maurer (Ct. Berne), Patrick Wasem (Ct. Schaffhouse), Patrick Porchet, Sylvain Kramer (Ct. Vaud). Les données analysées dans cet article ont été générées en collaboration avec le Genetic Diversity Centre (GDC), ETH Zurich. Les auteurs souhaitent remercier Silvia Kobel et Aria Minder pour leur précieux conseils lors de ces analyses. Cette étude a été co-financée par l’OFEV et le Centre Ecotox.
Avec l'abonnement en ligne, lisez le E-paper «AQUA & GAS» sur l'ordinateur, au téléphone et sur la tablette.
Avec l'abonnement en ligne, lisez le E-paper «Wasserspiegel» sur l'ordinateur, au téléphone et sur la tablette.




Kommentare (0)